
 Login/Register
Login/Register Supplier Login
Supplier Login









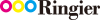
















Shengnan Fu, ⊥Fan C. Wang,⊥ Qiyuan Song, Zhihai Fan, Huaxiang Yang, Zhihua Huang,* Qiang Lu,* and Jun Chu*
Cite This: https://doi.org/10.1021/acsami.5c07451
Read online: Re-Understanding Silk Aggregates as Bioactive Agents | ACS Applied Materials & Interfaces
ABSTRACT: Bioactive agents play a pivotal role in determining the function and performance of biomedical products and devices across tissue engineering, regenerative medicine, and cosmetic applications. Silk fibroin (SF) is an extensively used structural
biomaterial, yet its inherent bioactive potential remains unexplored for material design. In this study, we developed β-sheet-rich silk nanofiberaggregates (BSNFs) with controlled diameters at ∼20 nm through structural bioinspiration from native silk fibers. We systematically compared our engineered BSNFs to two other distinct forms, conventional amorphous SF solution (ASF) and recombinant silk protein (RSF), to reveal the influence of the material conformation and nanostructure on their bioactivity. Notably, while ASF demonstrated better in vitro antioxidant capacity, both BSNFs and RSFs exhibited enhanced cellular antioxidant activity through improved phagocytic uptake. The nanofibrousstructure coupled with β-sheet-rich conformation facilitated cellular internalization and, thus, enhanced intracellular bioactivity. BSNFs outperformed ASF and RSF in stimulating cellular proliferation and migration, and they have better anti-inflammatory effects.Remarkably, BSNFs showed exceptional skin barrier protection capability and versatile activities in tissue repair and transepidermal delivery applications. This work establishes a paradigm shift by redefining SF aggregates as inherently bioactive components with their distinct nanostructures serving as critical activity modulators. Our findings not only reinforce silk’s value beyond structural applications but also provide a rational design for developing next-generation functional silk biomaterials with enhanced bioactivities.
KEYWORDS: silk fibroin,aggregates, bioactive agents, tissue regeneration, cosmetics
1. INTRODUCTION
Silk fibroin (SF) has been widely used as scaffolds in tissue regeneration and carriers in drug delivery due to its biocompatibility, tunable degradation, and ease of fabrication.1−3 To address diverse biomedical needs, various SF-based biomaterials such as scaffolds,hydrogels, films,coatings, and filaments have been engineered as structural matrices to support cell proliferation and tissue ingrowth.4−8 While considerable efforts have been devoted to tailoring SF’s hierarchical nano- and microstructures, anisotropic surface topographies, and mechanical properties to achieve specific biofunctions,9 its intrinsic potential as a bioactive agent for actively modulating cellular responses or therapeutic interventions remains under-explored. Natural SF filaments protect silkworm against environmental stressors such as sunlight, rainwater, etc., suggesting its inherent antioxidant capacities. Such bioactivity further corresponded to SF’s efficacyin promoting skin barrier restoration and mitigating skin allergic reactions.10,11 Revealing the structurally dependent bioactivity of SF could revolutionize its applications in functional biomaterials and drug delivery systems, opening new frontiers in biomedical research.
To harness the potential of SF as a bioactive agent, it is critical to identify the core factors governing its biological activity. Previous studies have demonstrated that SF’s physical characteristics, including mechanical properties, degradation kinetics, and hydrophilicity, can be precisely tuned by modulating its hierarchical assembly. 12−14 This suggests that controlling the assembly process of SFs may offera viable strategy to enhance its bioactivity. Conventionally, most SF-based biomaterials are usually made by dissolving silk fibers in concentrated salt solution and subsequently dialysis to facilitate their following fabrication; thus the amorphous SF (ASF) solutions serve as the foundational precursor for most SF-based materials.13,15,16 The ASF solutions were fabricated to form scaffolds,hydrogels, and films and then transformed into stable β-sheet-rich structures with after-treatments such as ethanol or methanol annealing.4,5,8 Recently, β-sheet-rich SF nanofibers (BSNFs) have been engineered through controlled assembly, which enabled the fabrication of SFs with tailored physical performances and biocompatibility. 17−19 Intriguingly, despite sharing similar β-sheet-rich secondary structures, ASF- and BSNF-derived materials exhibit markedly different behaviors in terms of degradation profiles, mechanical strength, and bioactivity,
highlighting the critical influence of higher-order assembly.20
Received: April 14, 2025
Revised: June 3, 2025
Accepted: June 4, 2025
--------------------------------------------------------------------------------------------------
Further expanding this paradigm, we have developed recombinant SF (RSF) through protein engineering to augment anti-
inflammatory capabilities. While ASF primarily adopts amorphous conformations, both RSF and BSNF exhibit β-sheet-rich
conformations, and such structural dichotomy makes these three SF variants (ASF, BSNF, and RSF) ideal model systems for
investigating how higher-order assembled organization dictates bioactivity.
Here, leveraging this structural dichotomy, we systematically investigated these three SF variants (ASF, BSNF, and RSF) to establish how their nanoscale assembly states govern bioactivity through comprehensive characterization of their secondary structures and higher-order organization. Their antioxidant capacities were evaluated through chemical redox assay and cellular antioxidant activity, which showed contradictory antioxidant results. These SFs had different phagocytotic activities that depend on the assembled states. The SF aggregates with high phagocytotic activity achieved significant better bioactivity such as antioxidant capacity, anti-inflammatory property, and better protective effects on the skin barrier under UVB treatment. Thus, the aggregate structures of SFs influenced the phagocytosis, which then determined their bioactivity.
2. RESULTS AND DISCUSSION
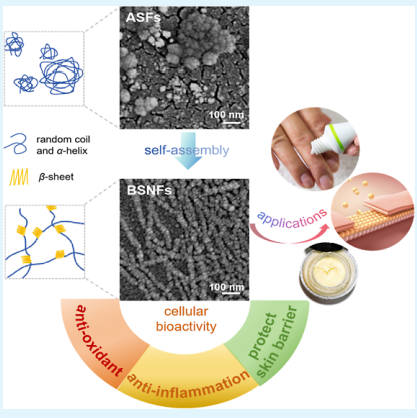
2.1. Morphology and Aggregates of Different SFs. Amorphous SFs (ASFs) remain hydrophilic for aqueous dissolution where the amorphous state spontaneously changes to a β-sheet structure in solution, resulting in hydrogel formation
or precipitation.21,22 Uncontrolled assembly led to inhomogeneous aggregates. As shown in Figure 1A,B, ASFs were composed of particles with a size of 10−1000 nm, confirming the wide size distribution. The concentration and temperature-
controlled assembly of SF in water developed stable water-dispersible BSNFs with diameter of ∼20 nm (Figure 1A) due to
their strong negative charge repulsion (Figure 1D). 23 Unlike ASF, BSNFs exhibited uniform nanofibrous structures with a
diameter of about 20 nm (Figure 1A). Similar to our previous study, 24 two size peaks appeared at about 20 and 100 nm due to
its nanofibrous structure (Figure 1B). FTIR and XRD curves revealed that ASF had typical random conformations, while BSNF exhibited β-sheet-rich structures (Figure 2). Despite sharing identical molecular origins and maintaining transparency below 0.5 wt % in aqueous solution, the two SFs had totally different aggregate structures (random vs β-sheet, inhomogeneous particles vs homogeneous nanofibers). Recently, RSFs were developed and used as bioactive agents in the cosmetic field. Although little study realized the aggregate structure of RSFs, the hydrogel state at an SF concentration of 2wt % similar to that of BSNFs implied the existence of aggregates with a hydrophobic crystal structure (Figure 1D). Scanning electron microscopy (SEM) images and size results showed nanofiberformation with diameters of about 40−60 nm (Figure 1A,B), significantly bigger than that of BSNFs. FTIR and XRD indicated a high content of β-sheet structure (Figure 2A,B). Compared to that of BSNF, a new small peak at 15.9° appeared in the XRD curve of RSF, but the β-sheet peak at 8.9° almost disappeared. The results suggested that RSFs contained both Silk I and Silk II conformations. The deconvolution of the amide I region from FTIR revealed different contents of the β-sheet in the three SFs (Figure 2C,D), where the highest content of the β-sheet was achieved by BSNF (46.1%), followed by RSF (36.0%) and ASF (17.6%) (Figure 2D). While high β-sheet content typically confers water insolubility due to increased hydro-phobicity, both BSNF and RSF maintained aqueous dispersibility through electrostatic stabilization, as evidenced by their strong negative zeta potentials (−40.5 ± 2.5 mV and −30.3 ± 0.7 mV, respectively; Figure 1C).25 Previous studies indicated that thelyophilized BSNFs with a β-sheet-rich structure failed to be re-dispersible in water due to their hydrophobic property.22 The BSNFs could maintain their conformations and aggregate structures after lyophilization treatment, favoring their application in SF scaffold materials. Repeated lyophilization treatment never changed the conformations of SF aggregates, which confirmed the stability (Figure S1). Although the lyophilized ASFs remained soluble in water due to their hydrophilic amorphous state, the lyophilized RSFs with a high content of β-sheet were insoluble in water, similar to the BSNFs (Figure 1E). Thus, the three SFs with similar molecular composition formed various aggregates in water, facilitating our understanding of the effectof the aggregate structure on bioactivities.
Figure 1. Structure and appearance of SFs: (A) SEM images; (B) particle size distribution; (C) zeta potential (n = 3, statistically significant***p ≤ 0.001 and ****p ≤ 0.0001); (D) appearance of SF aqueous solution and hydrogel (SF concentration 2 wt %); (E) redispersible behavior of different lyophilized SFs in water. The red arrows indicate the insoluble lyophilized BNSFs and RSFs. ASFs were aqueous SF molecules, BSNFs were β-sheet-rich SF nanofibers,and RSFs were recombinant SF aggregates.
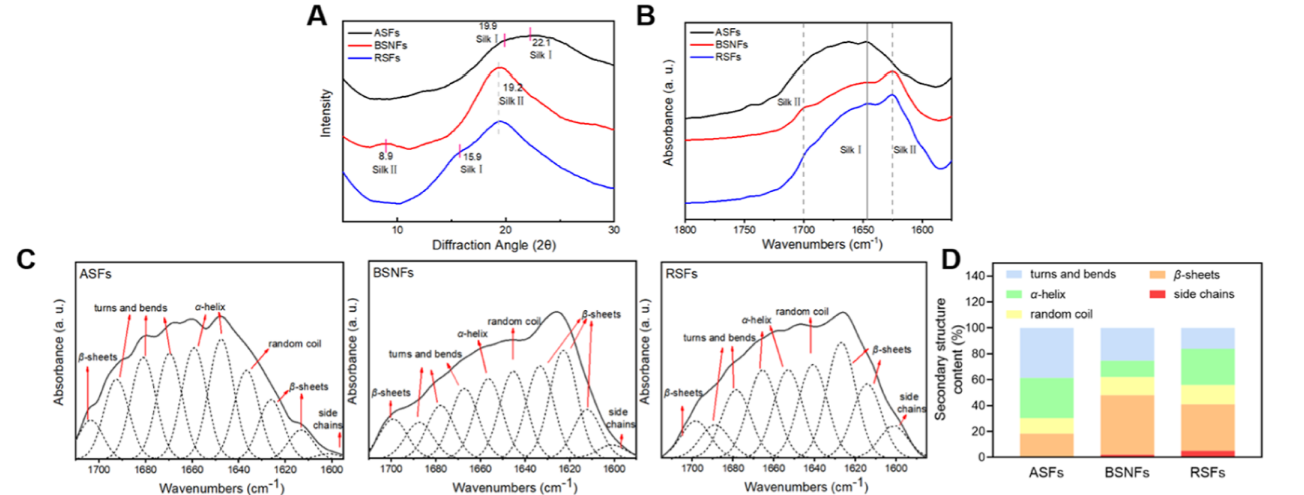
Figure 2. Conformational compositions of differentSFs: (A) X-ray diffraction curves of the SFs; (B) FTIR spectra of differentSFs; (C) deconvolution of the amide I region from FTIR; (D) ratios of secondary structures in differentSFs. ASFs were aqueous SF molecules, BSNFs were β-sheet-rich SF nanofibers,and RSFs were recombinant SF aggregates.
2.2. The Antioxidant Capacity of Different SFs. The antioxidant capacity of SF materials appeared to be mediated by the phenyl hydroxyl group of tyrosine. 26,27 A previous study indicated that the amorphous SF (ASF) in aqueous solution had
rich tyrosine on the surface to facilitate a horseradish peroxidase (HRP) cross-linking reaction.28 During SF assembly, these functional groups undergo spatial redistribution where the groups were buried inside the assemblies, rendering them inaccessible to HRP cross-linking.26 These structural differences between ASF and BSNFs suggested potential variations in their antioxidant activity. As shown in Figure 3A−C, chemical antioxidant assays (2,2′-azinobis(3-ethylbenzothiazoline-6-sul-fonic acid) (ABTS), 2,2-di(4-tert-octylphenyl)-1-picrylhydrazyl (DPPH), and hydroxyl radical scavenging) demonstrated superior activity for ASF compared to both BSNF and RSF at concentrations of 0.5−5 mg/mL, where RSFs had antioxidant activity similar to the BSNFs. Based on the antioxidant evaluation, it seems that ASFs have better bioactivity than the β-sheet-rich aggregates (BSNFs and RSFs). Surprisingly, in vitro cellular antioxidant assessment revealed an inverse trend. When L929 cells were treated with the three SFs for 24 h and then exposed to 1.6 mM H 2 O 2 for 1 h, BSNFs-treated cells showed 2.14-fold greater reactive oxygen species (ROS) clearance than ASFs-treated cells (Figure 3D,E). Notably, while RSFs and BSNFs showed comparable in vitro antioxidant capacities, BSNFs have demonstrated significantly better cellular ROS scavenging. These contrasting results suggest that chemical antioxidant capacity primarily reflectstyrosine accessibility, and cellular antioxidant behavior involves additional regulatory mechanisms beyond simple phenolic group availability where β-sheet-rich nanostructures (BSNF) may facilitate more efficient cellular uptake or intracellular processing. Several factors such as group distribution, nanomorphology, and mechanical property could influence both the antioxidant capacity and the cellular internalization of SF materials. Considering that these factors cannot be totally determined at present study, the underlying mechanism remains unclear. Although further study is required to explore the inherent mechanism of regulating antioxidant capacity of SF materials, the dichotomy between solution-phase and cellular antioxidant performance highlights the complex structure−activity relationships in SF-based systems, emphasizing the need for comprehensive evaluation when developing SF biomaterials for antioxidant applications.
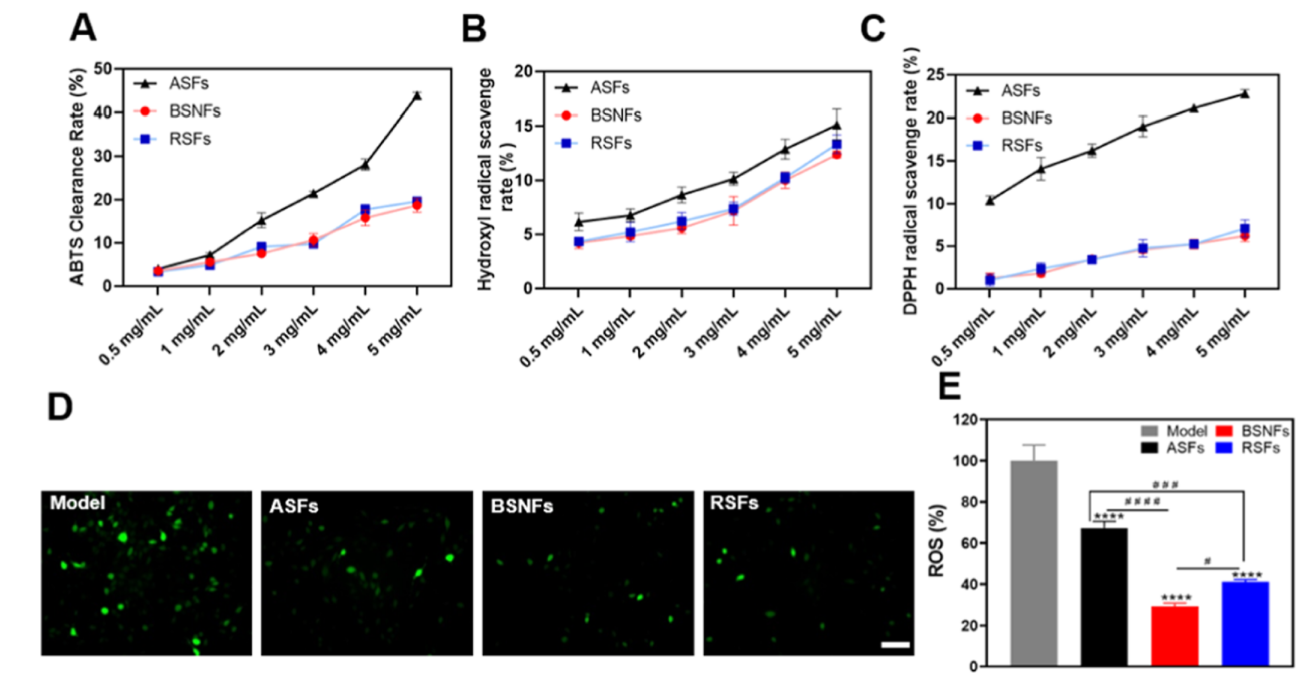
Figure 3. Antioxidant evaluation of differentSFs: Scavenging rates of (A) ABTS, (B) hydroxyl radicals, and (C) DPPH radicals; (D) the fluorescence photos of H 2 O 2 -stimulated L929 fibroblaststreated with differentSFs. The intercellular ROS was stained green. Scale bar = 100 μm; (E) relative ROS fluorescence intensity of different SF groups based on the model group. ASFs were aqueous SF molecules, BSNFs were β-sheet-rich SF nanofibers,and RSFs were recombinant SF aggregates. The model group received hydrogen peroxide treatment without prior material incubation. n = 3. Statistically significant****p ≤ 0.0001 vs the model group. Statistical significancewas determined by intergroup comparisons, #p ≤ 0.05, ###p ≤ 0.001 and ####p
≤ 0.0001.
2.3. Cellular Internalization Efficiency of Different SFs. Previous studies have established BSNFs as effective trans-
dermal drug carriers, where their nanoscale dimensions and hydrophobic properties facilitate skin penetration.
1,16 Considering the structural similarity between the cytomembrane and stratum corneum, we hypothesized that β-sheet-rich aggregates might similarly enhance cellular internalization and consequent bioactivity. Results of phagocytosis studies confirmed this hypothesis, demonstrating superior cellular uptake of BSNFs compared with both RSFs and ASFs, as quantified by flow
cytometry (Figure 4A) and visualized via confocal laser scanning microscopy (CLSM) (Figure 4B,C). The enhanced cellular
interaction of BSNFs correlated with their exceptional transdermal performance (Figure 4D,E) and cellular antioxidant
capacity. Notably, while both BSNFs and RSFs possess β-sheet rich structures, BSNFs’ smaller diameter (20 nm vs 40−60 nm
for RSFs) resulted in significantlygreater cellular internalization. These findings reveal that β-sheet organization promotes
cellular interaction through optimized hydrophobicity and nanostructure. The nanofiber diameter critically influences
phagocytic efficiency, and their cellular bioactivity can be enhanced by engineering these physical parameters. The length
is the more significant factor that influences intracellular behaviors. Our recent study revealed significant improved
cellular internalization through shortening the length of BSNFs.29
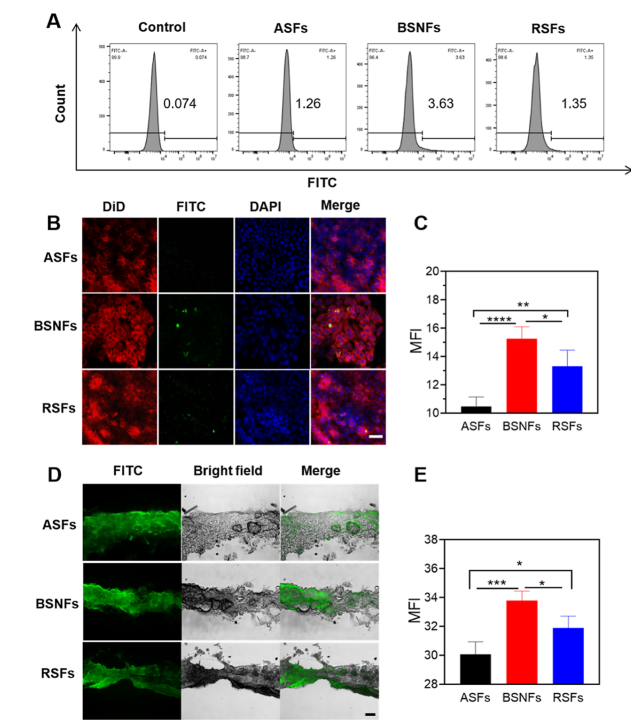
Figure 4. Bioabsorption behavior of differentSFs: (A) Cellular retention of FITC-labeled samples in HaCaT cells based on flow cytometry; the control group (negative control) was incubated in the SF material free medium. (B) Confocal laser scanning microscopy images of HaCaT cells cultured with ASFs, BSNFs, and RSFs for 24 h, Scale bar = 50 μm; DID-labeled cell membranes exhibited red fluorescence,FITC-labeled material showed green fluorescence,and DAPI-stained nuclei displayed blue fluorescence.(C) Mean fluorescence intensity (MFI) of FITC in (B); (D) fluorescent microscopy images of the dorsal skin of mice when differentSFs were applied for 24 h in vitro, Scale bar = 200 μm; the green fluorescence signature exclusively originated from FITC-conjugated material (E) MFI of FITC in (D). ASFs were aqueous SF molecules, BSNFs were β-sheet-rich SF nanofibers,and RSFs were recombinant SF aggregates. Data presented as mean ± SD, n = 3. Statistically significant*p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001.
Further enhancement of the phagocytic efficiency through controlling both the diameter and length of BSNFs is
anticipated, which will be realized in our future study. Zeta potential and conformation compositions could also influence
the phagocytic efficiency. High β-sheet content is considered to favor the adhesion of SF materials on the cells and then improve
the cellular internalization, which might be the other reason for better phagocytic efficiency of BSNFs. Higher negative charge
brought strong repulsion with the cells but also inhibited further aggregation. So, suitable Zeta potential of the SF aggregates
should facilitate the cytophagy. This structure−activity relationship suggests promising strategies for designing SF-based
delivery systems with tunable cellular interactions, where the controlled assembly of β-sheet structures can be leveraged to
optimize biological performance.
2.4. Cellular Bioactivities of Various SFs. Our previous studies have revealed the excellent stability of β-sheet-rich
aggregates in vitro and in vivo in scaffold and hydrogel systems. 3,7,13,15 Our present results also revealed that different
SF aggregates retained their original structures when cultured in the cellular medium for 24 h, suggesting its stability (Figure S2).
The β-sheet-rich aggregates as vehicles could be engulfed by thecells and then degraded inside the cells to deliver drugs.1,6,16,29 Thus, the different cellular internalizations should influence the bioactivity of the SF aggregates. The cellular bioactivities of different SF aggregates were evaluated by examining their effects on L929 cell proliferation and migration (Figure 5A−C). All SF-treated groups showed enhanced cell proliferation compared to the SF-free control, with β-sheet-rich aggregates (BSNF and RSF) demonstrating significantly greater stimulation than amorphous ASF. Notably, BSNF that exhibited the highest
cellular internalization promoted the stronSgest proliferative response. In scratch migration assays, BSNFs-treated cells
migrated 4.44 times faster than ASF-treated cells and 2.68 times faster than RSF-treated cells at 100 μg/mL, reaching 57.8% of
the migration rate observed with a 0.5% fetal bovine serum (FBS) positive control. This indicates that BSNFs not only
enhance proliferation but also actively promote wound healing processes.
Several studies also suggested the anti-inflammatory potential of SF materials.11,30 As expected, through their dose-dependent
modulation of cytokine secretion in macrophages, all SF variants stimulated IL-10 (anti-inflammatorycytokine) production while
suppressing IL-6 (proinflammatory cytokine) release compared to LPS controls, with BSNF showing the most potent effects
(Figure 5D,E). The optimized anti-inflammatory behaviors of BSNFs might be due to multiple factors such as better cellular
uptake efficiency, suitable nanostructure for macrophage polarization, and better biological interpretation and cellular
adhesion of the β-sheet-rich structure. Further studies will be continued to clarify the roles of various cues, facilitating the
design of bioactive SF materials in future. These results demonstrate that SF aggregates exhibit multiple bioactivities
including pro-proliferative, pro-migratory, and anti-inflammatory effects,all of which can be optimized by controlling their
assembly state. The superior performance of BSNFs highlights the crucial role of nanostructure-dependent cellular internal-
ization in enhancing bioactivity, reinforcing SF’s potential as a tunable bioactive agent for regenerative medicine applications.
The structure−activity relationships revealed in this study provide important guidance for designing SF-based biomaterials
with tailored biological functions.
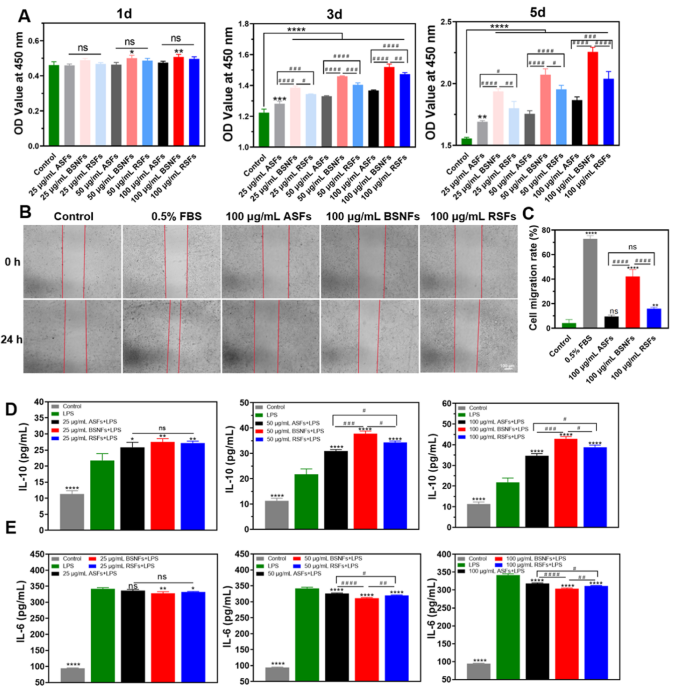
Figure 5. Cellular bioactivity of differentSFs: (A) Proliferation behavior of L929 fibroblastscultured with differentSFs; (B,C) migration behavior of L929 cultured with differentSFs, ns p > 0.05, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001 vs the control group. Statistical significance was determined by intergroup comparisons, #p ≤ 0.05, ##p ≤ 0.01, ###p ≤ 0.001, and ####p ≤ 0.0001; (D,E) secretion of IL-10 and IL-6 from RAW 264.7 cells cultured with various SFs. In the lipopolysaccharides (LPS) group, 200 ng/mL of LPS was added to the medium, and the cells were cultured for 8 h. Subsequently, the macrophages in the experimental group were cultured in medium containing differentSFs for 24 h after LPS treatment. The control group was the cells cultured in the LPS-free medium. ASFs, BSNFS, and RSFs were the LPS-treated cells cultured in the medium containing
ASFs, BSNFs, and RSFs, respectively. Data presented as mean ± SD, n = 3, ns p > 0.05, *p ≤ 0.05, **p ≤ 0.01, and ****p ≤ 0.0001 vs the LPS group.
2.5. Protective Effects of Different SFs. The protective effects of differentSF aggregates against UVB-induced damage
were systematically investigated. Building on silk fibers’natural role in protecting silkworms from ultraviolet radiation,31
we evaluated their potential to suppress cellular apoptosis. Using JC-1 fluorescence as a mitochondrial membrane potential
(MMP) indicator, we observed that UVB irradiation (50 mJ/cm 2 ) caused significantapoptosis in HaCaT cells,32
evidenced by the shift from red (healthy mitochondria) to green fluorescence (apoptotic cells) (Figure 6A). All SF variants
demonstrated protective effects, with β-sheet-rich aggregates showing a superior performance. Notably, BSNF treatment
maintained mitochondrial health comparable to untreated controls, while ASF and RSF provided intermediate protection,
suggested by measured MMP. Cell viability assays corroborated these findings(Figure 6B). UVB exposure reduced the viability
to 40.9% in unprotected cells, while SF treatment improved survival to 61.3% (ASF), 66.6% (RSF), and 87.1% (BSNF). The
enhanced protection by BSNFs with their smaller diameter (∼20 nm) suggests structure-dependent bioactivity.
Prolonged UVB exposure induces photoaging and injures skin barrier. 33,34 Claudin 1, filaggrin (FLG), and loricrin (LOR) are
typical proteins to repair skin barrier.31−33 We further examined SFs’ ability to mitigate UVB-induced photoaging by monitoring
key skin barrier proteins (Figure 6C−H). All three SF aggregates restored expression of Claudin1, FLG, and LOR that were
diminished by UVB exposure. BSNFs exhibited the most pronounced effect,increasing Claudin1 expression by 4.16-fold
compared to UVB-damaged controls and showing 2.82-fold and 1.25-fold improvements over ASF and RSF treatments,
respectively. Similar trends were observed for FLG (1.65-fold vs control) and LOR (1.26-fold vs control), with BSNFs
consistently outperforming other SF variants. These results have demonstrated that SFs preserved skin barrier function by
maintaining key structural proteins under UV irradiation, and the protection efficacy correlated with aggregate structure,
among which β-sheet-rich nanofibers show optimal performance. Such superior bioactivity of BSNFs likely stems from their
enhanced cellular internalization. The structure-dependent protective effectsobserved in this study highlight the potential
of engineered SF aggregates, particularly BSNFs, for photo-protection and skin barrier maintenance applications.
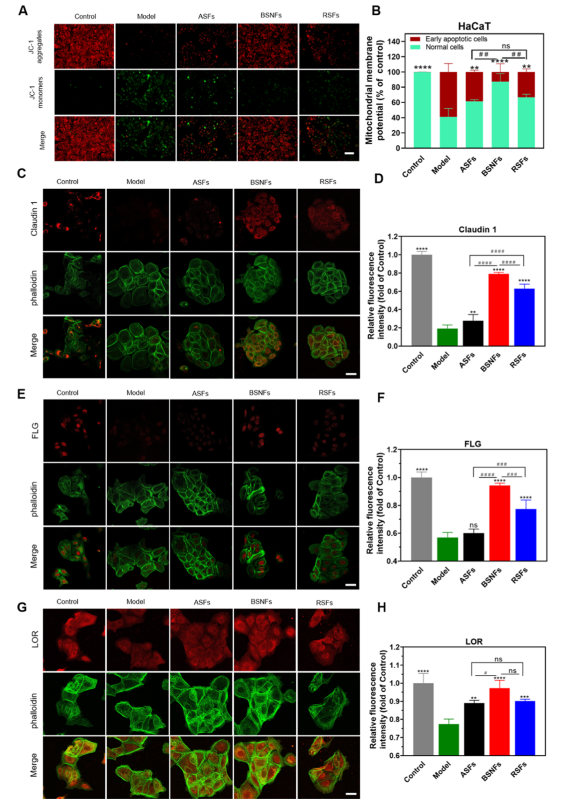
Figure 6. Protective effectsof differentSFs on UVB-exposed HaCaT cells: (A,B) Effectsof differentSFs on MMP (ΔΨ m ). HaCaT cells were exposed to 50 mJ/cm 2 UVB and then incubated with or without differentSFs for 24 h. MMP was evaluated by JC-1 assay; (C,D) representative images and fluorescence intensity of Claudin1-stained cells; (E,F) representative images and fluorescence intensity of FLG stained cells; (G,H) representative images and fluorescenceintensity of Loricrin LOR stained cells. Scale bar, 50 μm. ASFs, BSNFs, and RSFs were the UVB-treated cells cultured in the medium containing ASF, BSNFs, and RSFs, respectively. The control group was the cells without UVB irradiation, whereas the model group was the UVB-treated cells cultured in SF-free medium. Data presented as mean ± SD, n = 3, ns p > 0.05, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤
0.0001 vs the model group. Statistical significance was determined by intergroup comparisons, #p ≤ 0.05, ##p ≤ 0.01, ###p ≤ 0.001, and ####p ≤ 0.0001.
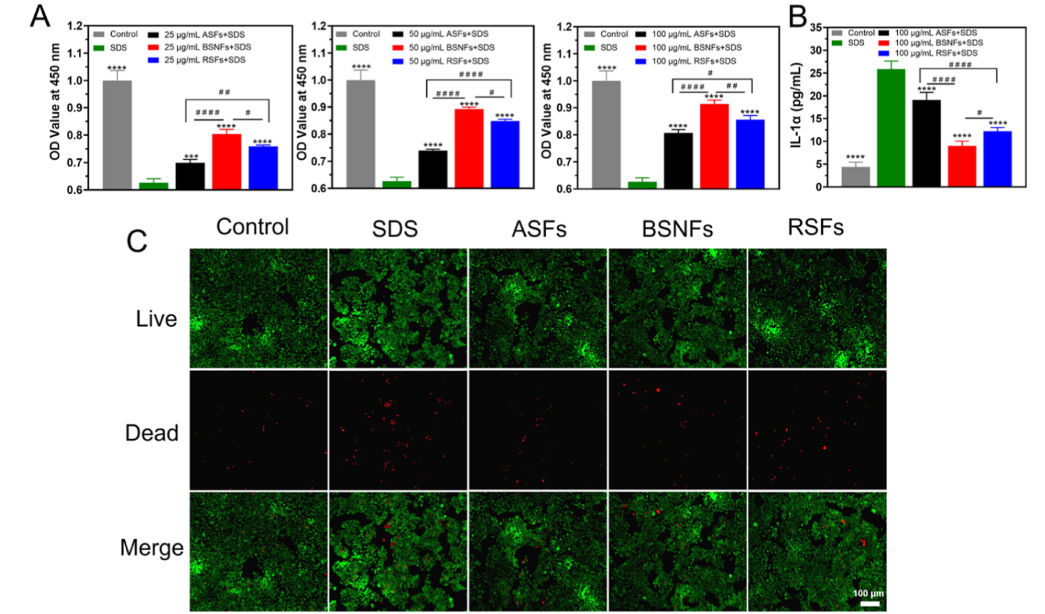
Figure 7. Barrier repair effects of SFs on SDS-induced HaCaT cell damage: (A) Cell viability of SDS-stimulated HaCaT cells when treated with differentSFs for 24 h. (SF concentration, 25, 50, and 100 μg/mL) n = 3; (B) IL-1α levels secretion from SDS-induced HaCaT cells cultured with different SFs for 24 h (SF concentration, 100 μg/mL) n = 4; (C) cellular viability quantified by live/dead assay (n = 3). The control group was the cells without SDS treatment, whereas the SDS group was the SDS-treated cells cultured in the SF-free medium. ASFs, BSNFs, and RSFs were the SDS-treated cells cultured in the medium containing ASFs, BSNFs, and RSFs, respectively. Data presented as mean ± SD. Statistically significant***p ≤ 0.001 and ****p ≤ 0.0001 vs the SDS group. Statistical significancewas determined by intergroup comparisons, #p ≤ 0.05, ##p ≤ 0.01, and ####p ≤
0.0001.
Beyond protection from UV damage, SF demonstrated significant protective effects against chemically induced skin
damage. We evaluated their efficacy against sodium dodecyl sulfate (SDS), a common surfactant in cosmetics and cleaning
products known to compromise skin barrier function. 35,36 Using an established SDS-induced cytotoxicity model, 37
we found that 10 μg/mL SDS reduced HaCaT cell viability to 62.6% (Figure 7A). All SF variants mitigated this damage in a dose-dependent manner, with β-sheet-rich BSNFs showing superior protection compared with both ASFs and RSFs. Although the dried BSNFs are difficult to develop various biomaterial forms such as hydrogel and scaffolds due to the poor redispersibility after
freeze-drying, the water insolubility of the dried BSNFs enabled the formation of a stable surface-adsorbed protective layer,
which would enhance the biological performance in cosmetics. The enhanced protective effectof BSNFs was further confirmed
through both live−dead staining assay and IL-1α secretion results (Figure 7B,C). These findings,combined with previously
demonstrated multifunctional properties (antioxidant capacity, anti-inflammatoryactivity, cell proliferation/migration enhance-
ment, and transdermal delivery potential), position SF aggregates, particularly BSNFs, as promising bioactive agents
for transdermal drug delivery, skin barrier repair, cosmetic applications, and other advanced biomedical applications
(Table 1).

3. CONCLUSIONS
This study changed the recognition of silk materials from an inert scaffold to a functional bioactive agent and realized the
critical roles of the aggregate structures in influencing multiple bioactivities (Table 1). Compared to traditional water-soluble
amorphous silk materials, β-sheet-rich nanofiber aggregates exhibit enhanced cellular uptake and transdermal penetration,
leading to superior bioactivity across multiple functional parameters. Specifically,these optimized nanostructures exhibit
potent antioxidant and anti-inflammatory property, significantly promote cell proliferation and migration, and provide excep-
tional protection against both UV radiation and chemical stressors. The multifunctional performance of these SF
aggregates highlights their strong potential for advanced cosmetic formulations, transdermal drug delivery systems, and
skin repair and protection applications. These insights fundamentally reshape our understanding of SF bioactivity,
establishing a clear structure−function relationship that will guide future design of silk-based bioactive materials. The
demonstrated ability to tune biological performance through controlled assembly of SF nanostructures opens new possibilities for developing biomaterials with tailored therapeutic effects.
4. EXPERIMENTAL SECTION
4.1. Preparation of ASF Aqueous Solutions. ASF was prepared according to the reported protocol. 38 Bombyx mori cocoons were boiled in 0.02 M Na 2 CO 3 for 30 min and washed with distilled water for above 5 times to remove sericin proteins. The degummed silk was dissolved in 9.3 M LiBr (Sigma-Aldrich, St. Louis, MO, USA) at 60 °C and then dialyzed in distilled water for 72 h. After centrifugation at 9000 rpm for 20 min at 4 °C, the ASF solution was obtained and stored at 4 °C for further use.
4.2. Preparation of RSF and BSNF. RSF was supplied by Givaudan (Silk-iCare, 2 wt % RSF). Silk-iCare has typical protein
sequence of GPGXX repetitive domains and polyalanine (Poly-A) motifs and exhibits a molecular weight of 139.9 kDa. BSNFs were prepared via a concentration-dilution process. The ASF solution was concentrated to above 20 wt % at 60 °C for above 36 h to form metastable nanoparticles. The concentrated solution was diluted to 0.5wt % with distilled water and incubated at 60 °C until gel formation. The BSNFs were assembled in the formed gels. 4.3. Characterization of Different SF Aggregates. The SEM
samples were prepared based on our previous studies. 18,19 The samples were diluted with distilled water to below 0.05%, and then, 5 μL of the diluted solution was deposited onto a clean silicon wafer. After being air-dried at room temperature, the samples were fixed on the table with conductive tape, sputtered with platinum, and observed using a SEM instrument (Hitachi, Regulus 8100, Tokyo, Japan) at 5 kV. The zeta potential and particle size of the silk nanocarriers were measured with a Zetasizer (Nano ZS, Malvern, Worcestershire, U.K.). For FTIR, the freeze-dried samples were measured with a Nicolet FTIR 5700 spectrometer (Thermo Scientific, FL, U.S.A.) at a resolution of 4 cm −1 and a wavenumber range from 400 to 4000 cm −1 . The infrared
spectra covering the amide I region were deconvoluted with PeakFit software to reveal the contents of various secondary structures. The XRD curves of the freeze-dried samples were obtained on an X-ray diffractometer(Nano ZS90, Malvern, Instruments, Malvern, U.K.) with scan speed of 5 deg/min and scan range of 5−60°. 4.4. Cell Culture. The human epidermal keratinocyte cell line (HaCaT), NCTC clone 929 (L929 fibroblasts), and Mouse mononuclear macrophage (Raw 264.7) were purchased from the Chinese Academy of Sciences (Shanghai, China). HaCaT cells were incubated in Dulbecco’s modified Eagle medium (DMEM, Gibco, MD, U.S.A.) with 10% FBS (Gemini, California, U.S.A.) and 1% penicillin−streptomycin (Invitrogen, NY, U.S.A.) at 37 °C in a humidified 5% CO2 incubator. L929 fibroblasts and Raw 264.7 cells were incubated in Roswell Park Memorial Institute (RPMI) 1640 (Gibco, BRL, MD, U.S.A.) with 10% fetal bovine serum and 1% penicillin−streptomycin at
37 °C in a humidified 5% CO 2 incubator. 4.5.
Antioxidant Capacity Test. The differentSF samples were diluted with distilled water to 0.5, 1, 2, 3, 4, and 5 mg/mL. The antioxidant capacity of these solutions was measured with the Hydroxyl Radical Antioxidant Capacity assay kit, the ABTS assay kit, and the DPPH assay kit according to the protocols. All of the kits were purchased from Shanghai Yuanye Bio-Technology Co., Ltd. (Shanghai, China). 4.6. Detection of ROS. Intracellular production of ROS was measured using cell-permeable fluorescentdyes, DCFH-DA (5-(and-6)-chloromethyl-20,70-dichlorodihydrofluorescein diacetate, acetyl ester). 39 L929 fibroblastswere treated with ASF, BSNF, and RSF (SF concentration, 100 μg/mL) for 24 h and washed with PBS 3 times. The cells were further incubated with H 2 O 2 (1.6 mmol/L) for 1 h and washed with PBS three times. The cells were incubated with DCFH-DA-diluted solution (5 μM) for 20 min at 37 °C. After washing with PBS three times, the stained cells were measured with a fluorescence microscope (CLSM, Olympus FV10 inverted microscope, Nagano, Japan) at the excitation and emission wavelengths of 480 and 530 nm, respectively. The fluorescence intensity of the cells was analyzed by ImageJ.
4.7. Cellular Uptake. DifferentSF samples were stained with Alexa Fluor 488 (FITC, Thermo Fisher, Waltham, MA, U.S.A.) according to the reported method.29 1 mg of FITC powder was dissolved in 1 mL of dimethyl sulfoxide (DMSO, Thermo Fisher, Waltham, MA, U.S.A.). The samples were mixed with the FITC solution at a 10:1 ratio and stained at 4 °C for 6 h, followed by dialysis in a membrane at room temperature in the dark for 48 h. HaCaT were inoculated in a 24-well plate at a density of 2 × 10 4 cells/well and then incubated in the medium containing FITC-stained SFs (SF concentration, 100 μg/mL) at 37 °C for 24 h. Then the cells were washed with PBS three times, fixed with 4% paraformaldehyde (PFA, Sigma-Aldrich, St. Louis, Missouri) for 10 min, and stained with 5 μM DiD (Beyotime, China) and DAPI staining solution (DAPI). The stained cells were imaged with
a CLSM (TCSP 8, Leica, Germany). The fluorescence intensity was analyzed by ImageJ. The cellular uptake was also analyzed
quantitatively with flow cytometry (FACS Calibur, BD Biosciences, San Jose, California, USA). HaCaT were inoculated in a 6-well plate at a density of 1 × 10 5 cells/well and then cultured in the medium containing FITC-stained SFs (SF concentration, 100 μg/mL) at 37 °C for 24 h. The cells were washed with PBS three times, digested with trypsin, and analyzed with flow cytometry.
4.8. Transdermal Permeation In Vitro. A vertical Franz diffusion cell (RYJ-6B, Shanghai Huanghai Pharmaceutical Inspection Instrument Co., Ltd.) with a diffusion area of 2.2 cm 2 was used to evaluate ex vivo transdermal permeation. All animal procedures were performed in accordance with the guidelines for Care and Use of Laboratory Animals of Soochow University and approved by the Animal Ethics Committee of the Soochow University (Approval number: 202309A0789). The dorsal skin of mice (C57BL/6J, female, 6−8 weeks) was collected and cleaned with tweezers to remove the fur. The skin was then fixed
between the donor and the receptor chambers. The receptor chambers were filled with PBS (pH 7.4). FITC-stained SF samples (1 mL, 5 mg/ mL) were added to the donor compartment and incubated at 37 °C for 24 h. Then the skin was collected and washed with PBS three times. After fixation,dehydration, and tissue embedding, the frozen section skin was observed under a fluorescence microscope. The FITC fluorescence was counted by ImageJ and analyzed with Prism. Each sample study was repeated three times.
4.9. CCK-8 Assay. L929 fibroblasts were inoculated in a 96-well plate at a density of 1 × 10 4 cells/well and then incubated in the
medium containing SFs (SF concentration, 25, 50, or 100 μg/mL) at 37 °C. At predetermined time intervals (1, 3, and 5 d), the cells were collected and washed with the culture medium for 3 times. The cells were treated with CCK-8 (Beyotime, Shanghai, China) for 1 h at 37 °C and measured at 450 nm using a plate reader (Biotek Synergy 4, Winooski, VT, USA).
4.10. Cell Migration Capacity. The cell migration behaviors were evaluated with the scratch method. 40 L929 fibroblastswere cultivated in 6-well plates until reaching 80% confluence. The scratches were prepared with a 200 μL pipet. Then the cells were cultured in RPMI 1640 containing SFs (SF concentration, 100 μg/mL) at 37 °C for 24 h. As the positive and negative control, the RPMI 1640 containing 0.5% FBS and the pure RPMI 1640 without the serum were also used to culture the cells. The healing progress of the cell scratches was observed and the percentage of the healed area between the cell scratches was calculated based on the following formula: cell scratch healing rate = 100% × [cell scratch breadth at the beginning (0 h) − cell scratch breadth after 24 h (24 h)]/cell scratch breadth at the beginning (0 h). The positive control was 0.5% FBS and the negative control was serum-free medium. Cells were cultured in the medium with 100 μg/mL of SFs for 24 h and then the migration rate measured.
4.11. IL-6 and IL-10 Secretion of Raw 264.7. Raw 264.7 cells were inoculated in a 24-well plate at a density of 2 × 10 4 cells/well for 24 h. Then the cells were polarized with LPS (200 ng/mL) for 8 h. The polarized macrophages were cultured in medium containing different SFs (SF concentration, 25, 50, or 100 μg/mL) for 24 h. The supernatant was collected and used to detect the secretion of the cytokines through Mouse IL-10 ELISA Kit and Mouse IL-6 ELISA Kit (Wuhan Fine Biotech Co., Ltd., Wuhan, China). The macrophages were also cultured in the medium without LPS as negative control and cultured in the medium with LPS as positive control.
4.12. MMP Measurements. HaCaT cells were cultured in a 24-well plate and cultured in medium containing differentSFs (100 μg/ mL) for 24 h. They were washed with PBS three times and then irradiated with UVB 50 mJ/cm 2 . After UVB treatment, DMEM containing 10% FBS and SFs (100 μg/mL) were added to the cells and cultured at 37 °C for 24 h. The cells were washed three times with DMEM and incubated with an MMP detection kit containing JC-1 (20 μg/mL, Beyotime Biotechnology Co., Ltd., Beijing, China) at 37 °C for 20 min. After the cells were further washed with PBS three times and covered with DMEM containing 10% FBS, the fluorescence images were observed with a fluorescencemicroscope (CLSM, Olympus FV10 inverted microscope, Nagano, Japan). The fluorescence intensity was statistically analyzed by ImageJ and Prism.
4.13. Immunofluorescence Analysis. HaCaT cells were cultured in a 24-well plate and cultured in a medium containing differentSFs (100 μg/mL) for 24 h. They were washed with PBS three times and then irradiated with UVB 50 mJ/cm 2 . After UVB treatment, DMEM containing 10% FBS and SFs (100 μg/mL) were added to the cells and cultured at 37 °C for 24 h. Then HaCaT cells were fixed with 4% PFA, treated with 0.1% TritonX-100, and sealed with 3% BSA-PBS. The sealed samples were incubated with the primary antibodies including Claudin1, FLG (Santa Cruz Biotechnology, Inc.) and LOR (Proteintech Group, Inc., USA) overnight at 4 °C and then incubated with secondary antibody (dilution ratio 1:200) in the dark for 2 h. DAPI and phalloidin were also used to stain the cells. The fluorescenceimages of the cells were observed with an inverted fluorescence microscope
(CLSM, Olympus FV10 inverted microscope, Nagano, Japan).
4.14. SDS-Induced Cell Toxicity Assay. HaCaT cells were inoculated in a 96-well plate at a density of 1 × 10 4 cells/well and then cultured in the medium containing SDS (10 μg/mL) and SFs (25, 50, and 100 μg/mL) for 24 h at 37 °C. Then the cells were washed three times with PBS and treated with CCK-8 for 1 h. The absorbance was measured at 450 nm using a plate reader (Biotek Synergy 4, Winooski, VT, USA). As the positive and negative control, the cells were also cultured in the medium with/without SDS for 24 h at 37 °C. The viability was measured with the CCk-8 assay. HaCaT cells were stained using a LIVE/DEAD Viability/Cytotoxicity Kit (Thermo, US) (100 μg/mL SFs) and observed under a fluorescence microscope (CLSM,
Olympus FV10 inverted microscope, Nagano, Japan). The supernatant was collected and used to detect the secretion of the cytokines through a Human IL-1α ELISA Kit (Multi Science Biotech CO, Ltd., Hangzhou, China).
4.15. Statistical Analysis. The mean ± standard deviation was used to describe the data. Graphpad Prism software was employed for data analysis. One-way AVOVA with Tukey’s multiple comparison tests was used to compare differences between groups by GraphPad Prism Software. ns p > 0.05, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001 were considered to be significant.
■ ASSOCIATED CONTENT
* sı Supporting Information
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acsami.5c07451.
FTIR spectra of different SF aggregates after repeated lyophilization treatment and SEM images of differentSF
aggregates when cultured in the cellular medium for 24 h (PDF)
■ AUTHOR INFORMATION
Corresponding Authors
Zhihua Huang − Guangzhou Eggshell Network Technology Co,
Ltd, Guangzhou 440101, People’s Republic of China;
Email: EddyHuang@homefacialpro.com
Qiang Lu − Department of Orthopedics, The Second Affiliated
Hospital of Soochow University, Soochow University, Suzhou
215000, People’s Republic of China; orcid.org/0000-
0003-4889-5299; Email: Lvqiang78@suda.edu.cn
Jun Chu − Department of Pediatric Surgery, Shanghai
Children’s Medical Center, Shanghai Jiao Tong University
School of Medicine, Shanghai 200127, People’s Republic of
China; Email: Chujun516@sina.com
Authors
Shengnan Fu − Department of Orthopedics, The Second
AffiliatedHospital of Soochow University, Soochow University,
Suzhou 215000, People’s Republic of China
Fan C. Wang − Future Food Laboratory, Innovation Center of
Yangtze River Delta, Zhejiang University, Jiashan 314100,
People’s Republic of China
Qiyuan Song − Department of Orthopedics, The Second
AffiliatedHospital of Soochow University, Soochow University,
Suzhou 215000, People’s Republic of China
Zhihai Fan − Department of Orthopedics, The Second Affiliated
Hospital of Soochow University, Soochow University, Suzhou
215000, People’s Republic of China
Huaxiang Yang − Department of Orthopedics, The Second
AffiliatedHospital of Soochow University, Soochow University,
Suzhou 215000, People’s Republic of China
Complete contact information is available at:
https://pubs.acs.org/10.1021/acsami.5c07451
Author Contributions
S.F. and F.W. contributed equally to the work. The manuscript was written through contributions of all authors. All authors
have given approval to the finalversion of the manuscript.
Notes
The authors declare no competing financial interest.
■ ACKNOWLEDGMENTS
The authors thank the National Nature Science Foundation of China (52273143). We also thank the Social Development
Program of Jiangsu Province (BE2023699) for support of this work.
■ REFERENCES
(1) Wu, H.; Liu, S.; Xiao, L.; Dong, X.; Lu, Q.; Kaplan, D. L. Injectable
and pH-Responsive Silk Nanofiber Hydrogels for Sustained Anticancer
Drug Delivery. ACS Appl. Mater. Interfaces 2016, 8 (27), 17118−17126.
(2) Xiao, L.; Lu, G.; Lu, Q.; Kaplan, D. L. Direct Formation of Silk
Nanoparticles for Drug Delivery. ACS Biomater. Sci. Eng. 2016, 2 (11),
2050−2057.
(3) Xu, G.; Xiao, L.; Guo, P.; Wang, Y.; Ke, S.; Lyu, G.; Ding, X.; Lu,
Q.; Kaplan, D. L. Silk Nanofiber Scaffolds with Multiple Angiogenic
Cues to Accelerate Wound Regeneration. ACS Biomater. Sci. Eng. 2023,
9 (10), 5813−5823.
(4) Zhou, Z.; Cui, J.; Wu, S.; Geng, Z.; Su, J. Silk fibroin-based
biomaterials for cartilage/osteochondral repair. Theranostics 2022, 12
(11), 5103−5124.
(5) Gholipourmalekabadi, M.; Sapru, S.; Samadikuchaksaraei, A.;
Reis, R. L.; Kaplan, D. L.; Kundu, S. C. Silk fibroin for skin injury repair:
Where do things stand? Adv. Drug Delivery Rev. 2020, 153, 28−53.
(6) Yang, C.; Li, S.; Huang, X.; Chen, X.; Shan, H.; Chen, X.; Tao, L.;
Zhang, M. Silk Fibroin Hydrogels Could Be Therapeutic Biomaterials
for Neurological Diseases. Oxid. Med. Cell. Longevity 2022, 2022,
2076680.
(7) Cheng, W.; Ding, Z.; Zheng, X.; Lu, Q.; Kong, X.; Zhou, X.; Lu, G.;
Kaplan, D. L. Injectable hydrogel systems with multiple biophysical and
biochemical cues for bone regeneration. Biomater. Sci. 2020, 8 (9),
2537−2548.
(8) Ding, Z.; Cheng, W.; Mia, M. S.; Lu, Q. Silk Biomaterials for Bone
Tissue Engineering. Macromol. Biosci. 2021, 21 (8), No. e2100153.
(9) Yao, X.; Zou, S.; Fan, S.; Niu, Q.; Zhang, Y. Bioinspired silk fibroin
materials: From silk building blocks extraction and reconstruction to
advanced biomedical applications. Mater. Today Bio 2022, 16, 100381.
(10) Farokhi, M.; Mottaghitalab, F.; Fatahi, Y.; Khademhosseini, A.;
Kaplan, D. L. Overview of Silk Fibroin Use in Wound Dressings. Trends
Biotechnol. 2018, 36 (9), 907−922.
(11) Mazurek, Ł.; Szudzik, M.; Rybka, M.; Konop, M. Silk Fibroin
Biomaterials and Their Beneficial Role in Skin Wound Healing.
Biomolecules 2022, 12 (12), 1852.
(12) Rockwood, D. N.; Preda, R. C.; Yucel, T.; Wang, X.; Lovett, M.
L.; Kaplan, D. L. Materials fabrication from Bombyx mori silk fibroin.
Nat. Protoc. 2011, 6 (10), 1612−1631.
(13) Lu, Q.; Zhang, B.; Li, M.; Zuo, B.; Kaplan, D. L.; Huang, Y.; Zhu,
H. Degradation mechanism and control of silk fibroin. Biomacromo-
lecules 2011, 12 (4), 1080−1086.
(14) Fan, Z.; Xiao, L.; Lu, G.; Ding, Z.; Lu, Q. Water-insoluble
amorphous silk fibroin scaffolds from aqueous solutions. J. Biomed.
Mater. Res., Part B 2020, 108 (3), 798−808.
(15) Zhang, X.; Xiao, L.; Ding, Z.; Lu, Q.; Kaplan, D. L. Engineered
Tough Silk Hydrogels through Assembling beta-Sheet Rich Nanofibers
Based on a Solvent Replacement Strategy. ACS Nano 2022, 16 (7),
10209−10218.
(16) Wang, X.; Liu, K.; Fu, S.; Wu, X.; Xiao, L.; Yang, Y.; Zhang, Z.;
Lu, Q. Silk Nanocarrier with Tunable Size to Improve Transdermal
Capacity for Hydrophilic and Hydrophobic Drugs. ACS Appl. Bio
Mater. 2023, 6 (1), 74−82.
(17) Wang, H. Y.; Zhang, Y. Q.; Wei, Z. G. Dissolution and processing
of silk fibroin for materials science. Crit. Rev. Biotechnol. 2021, 41 (3),
406−424.
(18) Hou, J.; Ding, Z.; Zheng, X.; Shen, Y.; Lu, Q.; Kaplan, D. L.
Tough Porous Silk Nanofiber-Derived Cryogels with Osteogenic and
Angiogenic Capacity for Bone Repair. Adv. Healthcare Mater. 2023, 12
(17), No. e2203050.
(19) Ding, Z.; Lu, G.; Cheng, W.; Xu, G.; Zuo, B.; Lu, Q.; Kaplan, D. L.
Tough Anisotropic Silk Nanofiber Hydrogels with Osteoinductive
Capacity. ACS Biomater. Sci. Eng. 2020, 6 (4), 2357−2367.
(20) Sun, W.; Gregory, D. A.; Tomeh, M. A.; Zhao, X. Silk Fibroin as a
Functional Biomaterial for Tissue Engineering. Int. J. Mol. Sci. 2021, 22
(3), 1499.
(21) Kong, N. Self-Assembly of Bombyx mori Silk Fibroin. Methods
Mol. Biol. 2021, 2347, 69−82.
(22) Xu, L.; Wu, C.; Lay Yap, P.; Losic, D.; Zhu, J.; Yang, Y.; Qiao, S.;
Ma, L.; Zhang, Y.; Wang, H. Recent advances of silk fibroin materials:
From molecular modification and matrix enhancement to possible
encapsulation-related functional food applications. Food Chem. 2024,
438, 137964.
(23) Bai, S.; Liu, S.; Zhang, C.; Xu, W.; Lu, Q.; Han, H.; Kaplan, D. L.;
Zhu, H. Controllable transition of silk fibroin nanostructures: an insight
into in vitro silk self-assembly process. Acta Biomater. 2013, 9 (8),
7806−7813.
(24) Lu, Q.; Wang, X.; Lu, S.; Li, M.; Kaplan, D. L.; Zhu, H.
Nanofibrous architecture of silk fibroin scaffolds prepared with a mild
self-assembly process. Biomaterials 2011, 32 (4), 1059−1067.
(25) Zhang, X.; Zhang, Z.; Xiao, L.; Ding, Z.; He, J.; Lu, G.; Lu, Q.;
Kaplan, D. L. Natural Nanofiber Shuttles for Transporting Hydro-
phobic Cargo into Aqueous Solutions. Biomacromolecules 2020, 21 (2),
1022−1030.
(26) Partlow, B. P.; Bagheri, M.; Harden, J. L.; Kaplan, D. L. Tyrosine
Templating in the Self-Assembly and Crystallization of Silk Fibroin.
Biomacromolecules 2016, 17 (11), 3570−3579.
(27) Yang, Y.; Shao, Z.; Chen, X.; Zhou, P. Optical spectroscopy to
investigate the structure of regenerated Bombyx mori silk fibroin in
solution. Biomacromolecules 2004, 5 (3), 773−779.
(28) Partlow, B. P.; Hanna, C. W.; Rnjak-Kovacina, J.; Moreau, J. E.;
Applegate, M. B.; Burke, K. A.; Marelli, B.; Mitropoulos, A. N.;
Omenetto, F. G.; Kaplan, D. L. Highly tunable elastomeric silk
biomaterials. Adv. Funct. Mater. 2014, 24 (29), 4615−4624.
(29) Xiao, L.; Ding, Z.; Zhang, X.; Wang, X.; Lu, Q.; Kaplan, D. L. Silk
Nanocarrier Size Optimization for Enhanced Tumor Cell Penetration
and Cytotoxicity In Vitro. ACS Biomater. Sci. Eng. 2022, 8 (1), 140−
150.
(30) Gonzalez-Restrepo, D.; Zuluaga-Velez, A.; Orozco, L. M.;
Sepulveda-Arias, J. C. Silk fibroin-based dressings with antibacterial and
anti-inflammatory properties. Eur. J. Pharm. Sci. 2024, 195, 106710.
(31) Santos, J. H.; Hunakova, L.; Chen, Y.; Bortner, C.; Van Houten,
B. Cell sorting experiments link persistent mitochondrial DNA damage
with loss of mitochondrial membrane potential and apoptotic cell
death. J. Biol. Chem. 2003, 278 (3), 1728−1734.
(32) Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.;
Tzur, A. JC-1 alternative excitation wavelengths facilitate mitochondrial
membrane potential cytometry. Cell Death Dis. 2012, 3 (11), No. e430.
(33) Krutmann, J.; Berneburg, M. [Sun-damaged skin (photoaging):
what is new?]. Hautarzt 2021, 72 (1), 2−5.
(34) Wang, Z.; Chen, H.; Wang, Y.; Wu, C.; Ye, T.; Xia, H.; Huang, R.;
Deng, J.; Li, Z.; Huang, Y.; Yang, Y. Recombinant filaggrin-2 improves
skin barrier function and attenuates ultraviolet B (UVB) irradiation-
induced epidermal barrier disruption. Int. J. Biol. Macromol. 2024, 281
(Pt 1), 136064.
(35) Mizutani, T.; Mori, R.; Hirayama, M.; Sagawa, Y.; Shimizu, K.;
Okano, Y.; Masaki, H. Sodium Lauryl Sulfate Stimulates the Generation
of Reactive Oxygen Species through Interactions with Cell Membranes.
J. Oleo Sci. 2016, 65 (12), 993−1001.
(36) Petushkova, N. A.; Rusanov, A. L.; Zgoda, V. G.; Pyatnitskiy, M.
A.; Larina, O. V.; Nakhod, K. V.; Luzgina, N. G.; Lisitsa, A. V.
[Proteome of the human HaCaT keratinocytes: Identification of the
oxidative stress proteins after sodium dodecyl sulpfate exposur]. Mol.
Biol. 2017, 51 (5), 748−758.
(37) Rusanov, A. L.; Luzgina, N. G.; Lisitsa, A. V. Sodium Dodecyl
Sulfate Cytotoxicity towards HaCaT Keratinocytes: Comparative
Analysis of Methods for Evaluation of Cell Viability. Bull. Exp. Biol.
Med. 2017, 163 (2), 284−288.
(38) Tao, X.; Jiang, F.; Cheng, K.; Qi, Z.; Yadavalli, V. K.; Lu, S.
Synthesis of pH and Glucose Responsive Silk Fibroin Hydrogels. Int. J.
Mol. Sci. 2021, 22 (13), 7107.
(39) Li, Z.; Jiang, R.; Wang, M.; Zhai, L.; Liu, J.; Xu, X.; Sun, L.; Zhao,
D. Ginsenosides repair UVB-induced skin barrier damage in BALB/c
hairless mice and HaCaT keratinocytes. J. Ginseng Res. 2022, 46 (1),
115−125.
(40) Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.;
Falanga, V. Research Techniques Made Simple: Analysis of Collective
Cell Migration Using the Wound Healing Assay. J. Invest. Dermatol.
2017, 137 (2), e11−e16.